The cashew apple is a juicy and sweet fruit that loses its nutritional quality during the first week after falling from the cashew tree. This degradation is strongly linked to microorganisms that may present beneficial biotechnological potential. Therefore, the objective of this study is to characterize lactic acid bacteria and fermentative yeasts from cashew apples. Cashew apples collected from three localities (Tioro, Morovine, and Waraniene) in the Korhogo department (Northern Côte d’Ivoire) were washed in the laboratory, crushed, and subjected to fermentation. Samples were taken every two days over one week. Morphological studies focused on macroscopic and microscopic analyses of the strains. Physiological and biochemical tests of lactic acid bacteria and yeast isolates were all carried out in tubes. Molecular typing of these isolates was performed using the restriction enzymes DDeI or HinfI, and HaeIII. Characterization of lactic isolates revealed that all lactic acid bacteria isolated were bacillary in shape with catalase activity. They are homofermentative, mesophilic, acidophilic, and do not tolerate salinity beyond 4%. Moreover, they fermented lactose, glucose, maltose, fructose, starch, sucrose, and cellulose. They also produced exopolysaccharides. At the molecular level, all bands were observed at 1500 base pairs. Furthermore, the biochemical profile and enzymatic digestion performed with restriction enzymes identified the genus Lactobacillus, specifically Lactobacillus plantarum as the presumptive species. Characterization revealed yeasts of oval, elongated shape with lateral budding. Sediments were observed in their growth broths. They degraded urea and produced acetoin. They also fermented various sugars, except cellulose, citrate, and mannitol. All bands were observed at 500 base pairs. Biochemical tests and PCR confirmed that all isolates belonged to the genus Saccharomyces sp. Digestion with restriction enzymes (HaeIII and HinfI) divided them into two groups of Saccharomyces, of which 93.75% were Saccharomyces cerevisiae and 6.25% were Saccharomyces uvarum. The use of these presumptive species could serve in the production of cashew apple wine.
| Published in | International Journal of Microbiology and Biotechnology (Volume 11, Issue 1) |
| DOI | 10.11648/j.ijmb.20261101.12 |
| Page(s) | 9-19 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Cashew Apple, Fermentation, Lactic Acid Bacteria, Yeast, Enzymatic Digestion
Amplification program | Lactic acid bacteria | Yeasts |
|---|---|---|
Initial denaturation | 95°C / 4 min | 95°C/3 min |
Denaturation | 95°C / 60 s | 95°C / 30 s |
Annealing | 57°C / 60 s | 55°C / 60 s |
Extension | 72°C / 1 min | 72°C / 1 min |
Final extension | 72°C / 5 min | 72°C / 10 min |
TiJ | Yellow Apple from Tioro |
TiR | Red Apple from Tioro |
MoJ | Yellow Apple from Morovine |
MoR | Red Apple from Morovine |
PCR | Polymerase Chain Reaction |
PCA | Plate Count Agar |
OGA | Oxytetracycline-Glucose-Agar |
MRS | De Man, Rogosa, and Sharpe |
VP | Voges-Proskauer |
dNTP | Deoxynucleoside Triphosphates |
RFLP | Restriction Fragment Length Polymorphism |
YPD | Yeast Peptone Dextrose |
EDTA | Ethylenediaminetetraacetic Acid |
SDS | Sodium Dodecyl Sulfate |
DNA | Deoxyribonucleic Acid |
ADH | Arginine Dihydrolase Activity |
EtBr | Ethidium Bromide |
bp | Base Pairs |
| [1] | Kalui, C. M., Mathara, J. M., Kutima, P. M. Probiotic potential of spontaneously fermented cereal-based foods – a review. African Journal of Biotechnology; 2010, 9(17), 2490–2498. |
| [2] | Bourdichon, F., Casaregola, S., Farrokh, C., Frisvad, J. C., Gerds, M. L., Hammes, W. P., Harnett, J., Huys, G., Laulund, S., Ouwehand, A., Powell, I. B., Prajapati, J. B., Seto, Y. T., Schure, E., Van Boven, A., Vankerckhoven, V., Zgoda, A., Tuijtelaars, S., Hansen, E. B. Food fermentations: Microorganisms with technologically beneficial use. International Journal of Food Microbiology; 2012, 154, 87–97. |
| [3] |
ASTEP. Charter for school support staff: Interventions in school settings, 2015. Available at:
http://espe.univ-lorraine.fr/lamap/sites/espe.univlorraine.fr.lamap/files/ressources/1516_astep_charte_new.pdf [Accessed 10 Feb. 2024]. |
| [4] | Jay, J. M., Loessner, M. J., Golden, D. A. Modern food microbiology, 7th edition. Springer; 2005, USA, 39–57. |
| [5] | Adriana, J., Potus, J., Poiffait, A., Dauvillier, P. Introduction to nutritional analysis of foodstuffs. Tech. and Doc. Lavoisier, Paris; 1998, 254 p. |
| [6] | Couto, S. R., Sanromán, M. Á. Application of solid-state fermentation to the food industry—A review. Journal of Food Engineering; 2006, 76(3), 291–302. |
| [7] | Guiraud, J. P. Food microbiology. Dunod, Paris; 1998, 310–321. |
| [8] | Jean, M. E., Didier, B., Patrick, P., Caroline, G., Frederic, B., Jean-Charles R. Impact of fermentation processes on the antioxidant and immunomodulatory properties of Guiana and Forastero cocoa from French Guiana. Foods; 2021, 10, 522. |
| [9] | Idoui, T., Boudjerda, J., Leghouchi, E., Karam, N. E. Lactic acid bacteria from sheep’s Dhan, a traditional butter from sheep’s milk: Isolation, identification and major technological traits. Gr. Y. Aceites; 2009, 60(2), 177–183. |
| [10] | Badis, A., Guetarni, D., Moussa, B. B., Henni, D. E., Kihal, M. Identification and technological properties of lactic acid bacteria isolated from raw goat milk of four Algerian breeds. Food Microbiology; 2004, 21(5), 579–588. |
| [11] | Marconi, E., Sorrentino, E., Mastrocola, E., Coppola, R. Rapid detection of diaminopimelic acid in lactic acid bacteria by microwave cell wall hydrolysis. J. Agric. Chem.; 2000, 48, 3348–3351. |
| [12] | Joffine, J. N., Leyral, G. Microbiologie technique, Tome 1: Dictionnaire des techniques, 4th edition. CRDP d’Aquitaine, Paris; 2006. |
| [13] | Bouix, M., Leveau, J. Y. Analysis and control techniques in the food industry, 2nd edition. Tec & Doc, Lavoisier, Paris; 1991, 206–229. |
| [14] | Leveau, J. Y., Bouix, M., De Roissart, H. B. Lactic flora: analysis and control techniques in the food industry, 2nd edition. Tec & Doc, Lavoisier, Paris; 1991, 2–40. |
| [15] | Dellaglio, F., De Roissart, H., Torriani, S., Curk, M. C., Janssens, D. General characteristics of lactic acid bacteria. In Lactic Acid Bacteria, Uriage Lorica; 1994, 25–116. |
| [16] | Garvie, E. I. Genus Pediococcus. In Bergey’s Manual of Systematic Bacteriology, Springer New York; 1986, 2, 1075–1079. |
| [17] | Mayeux, J. V., Sandine, W. W. F., Elliker, P. R. A selective medium for detecting Leuconostoc organisms in mixed starter cultures. J. Dairy Sci.; 1962, 45, 655–656. |
| [18] | Branger, A., Richer, M. M., Roustel, S. Microchemistry and food. Educagri Edition, Paris; 2007. |
| [19] | El Sheikha, A. F., Métayer, I., Montet, D. A biological barcode for determining the geographical origin of fruit using 28S rDNA fingerprinting of fungal communities by PCR-DGGE: an application to Physalis fruits from Egypt. Food Biotechnology; 2010, 25(2), 115–129. |
| [20] | Ahmed, H. M., Gang, W., Hao, Z. Morphological and molecular tools for identification of Saccharomyces boulardii isolated from active dry yeast. International Journal of Agriculture Innovations and Research; 2019, 7(4), 2319–1473. |
| [21] | Kahina, M. M. Characterization of a bacteriocin produced by a lactic acid bacterium Leuconostoc pseudomesenteroides isolated from boza. Doctoral Thesis, Bacteriology, Pierre and Marie Curie University, Paris VI; 2011, 124 p. |
| [22] | Nang, N. N. K., Somsay, P., Kridsada, U., Apinun, K., Chalermpong, S., Saisamorn, L., Kalidas, S., Chartchai, K. Valorization of cashew apple waste into a low-alcohol, healthy drink using a co-culture of Cyberlindnera rhodanensis DK and Lactobacillus pentosus A14-6. Foods; 2025, 13(10), 1–24. |
| [23] | Oyetayo, A. M., Bada, S. O., Ajongbolo, K. Effect of fermentation on the proximate composition of cashew apple residue. International Journal of Interdisciplinary Innovative Research & Development; 2017, 2(1), 43–47. |
| [24] | Estefânia, F. G. Identification of lactic acid bacteria in fruit pulp processing byproducts and potential probiotic properties of selected Lactobacillus strains. Frontiers in Microbiology; 2016, 7(1371), 1–11. |
| [25] | Madalyd, Y. V. P., Wendy, L., Rodriguez, R. Effect of pH on the growth of three lactic acid bacteria strains isolated from sour cream. Universitas Scientiarum; 2020, 25(2), 341–358. |
| [26] | Martha, W. K., Joseph, W. M., John, M. N. Physicochemical, antioxidant, and sensory properties of functional mango (Magifera indica L.) leather fermented by lactic acid bacteria. Journal of Agriculture and Food Research; 2021, 6, 1–9. |
| [27] | Bekhouche, F. Lactic acid bacteria from raw cow’s milk and pectinolytic microorganisms from black and green olives: isolation and biochemical identification; evaluation and optimization of polygalacturonase production. Doctoral Thesis, University of Mentouri Constantine; 2006, 149 p. |
| [28] | Mechai, A. Isolation, characterization, and purification of bateriocins produced by indigenous lactic acid bacteria: physiological and biochemical studies. University of Badji-Mokhtar Annaba; 2009, 160 p. |
| [29] |
Kamar, A. Lactic acid bacteria isolated from Moroccan traditional foods: bacteriocin production and potential applications against multi-drug-resistant pathogens. Analytical Chemistry. University of Strasbourg; Sidi Mohamed Ben Abdellah University (Fes, Morocco); 2023, 108 p.
https://publication-theses.unistra.fr/public/theses_doctorat/2019/AMENSAG_Kamar_2019_ED222.pdf |
| [30] | Holzapfel, W. H. Appropriate starter culture technologies for small-scale fermentation in developing countries. International Journal of Food Microbiology; 2002, 75(3), 197–212. |
| [31] | Yaya, A. O., Doudjo, S., Kouakou, R. F., Ahou, C. I. Y., Ismael, A. C., Dogbo, M. A., Adolphe, Z., Emmanuel, A., Kouassi, B. Y. Isolation and characterization of indigenous Acetobacter strains from cashew apple and their potential use in vinegar production. Fermentation; 2025, 11(1), 1–18. |
| [32] | Pramod, K. S., Renu, S., Manoj, S., Sibanada, D., Vellaichamy, M., Laccy, P., Bharti, R. Microbial production of α-amylase from agro-waste: An approach towards biorefinery and bio-economy. Energy Nexus; 2024, 14, 1–14. |
| [33] | Yingying, L., Luyao, X., Juanjuan, T., Xiaomeng, W., Xueliang, Z., Yong, C., Wei, L. Structural characterization, rheological properties, and protection against oxidative damage of an exopolysaccharide from Leuconostoc citreum 1.2461 fermented in soy whey. Foods; 2016, 11(15), 2283. |
| [34] | Meryem, M. Isolation and identification of lactic acid bacteria and study of their antagonistic effect against pathogenic microorganisms. Thesis in Biological Sciences, University 8 May 1945–Guelma, Algeria; 2016, 1–159. |
| [35] | Houali, K. Identification of lactic acid bacteria isolated from raw milk (Annaba region), study of their technological capabilities and antagonistic effect of Lactobacillus on Staphylococcus sp., Pseudomonas aeruginosa, Escherichia coli, Saccharomyces cerevisiae, and Aspergillus clavatus. Doctoral Thesis, University of Badji Mokhtar, Annaba; 1999, 261 p. |
| [36] | Bourgeois, C. M., Leveau, J. Y. Analysis and control techniques in the food industry, Vol. III: Microbiological control. Lavoisier, Paris; 1991, 451 p. |
| [37] | Leveau, J. Y., Bouix, M. Industrial microbiology: microorganisms of industrial interest. Coll. Sciences and techniques agro-foods, 1st edition, Tec & Doc Lavoisier; 1993. |
| [38] | Bouchibane, M. Identification of lactic acid bacteria isolated from artisanal dairy products: technological abilities and trials for producing fermented milk. Doctoral Thesis, University Abdelhamid Ibn Badis – Mostaganem; 2023, 100 p. |
| [39] | David, O., Nestor, D. G., Santiago, C., Bernadette, K. C. Fermentation of a mango beverage with Lactiplantibacillus plantarum LP01: from bench to semi-industrial scale. Journal of Food; 2025, 23(1), 1–10. |
| [40] | Dennia, P. A. T., Jessica, P. B., Eliane, M. F. M., Patrícia, A. S. T. Isolation and identification of lactic acid bacteria in fruit processing residues from the Brazilian Cerrado and its probiotic potential. Food Bioscience; 2022, 48(2), 215–232. |
| [41] | Aspasia, A. N., Dimitra, D., Maria, E. F., Ellie, D., Petros, F., Chryssoula, T., Georgios, B. Genetic and technological characterization of vineyard- and winery-associated lactic acid bacteria. BioMed Research International, Article ID 508254; 2014, 8 p. |
| [42] | Irena, V., Marta, H., Blanka, J., Zdena, P. The morphology of Saccharomyces cerevisiae colonies is affected by cell adhesion and the budding pattern. Research in Microbiology; 2005, 15(6), 921–931. |
| [43] | Fumie, W. S., Youji, N., Munekazu, K., Masashi, H., Tohru, O. Influence of wine components on pellicle formation by pellicle-forming yeasts. OENO One; 2021, 55(3), 621–643. |
| [44] | Rani, P., Sharma, S., Garg, F. C., Raj, K., Wati, L. Ethanol production from potato flour by Saccharomyces cerevisiae. Indian Journal of Science and Technology; 2010, 3, 733–736. |
| [45] | Kurtzmann, C. P., Fell, J. W., Boekhout, T. The yeasts, a taxonomic study, 5th edition. Elsevier, Amsterdam; 2011, 3–9. |
| [46] | Paulina, J., Jun-Yong, C., Eckhard, B., Mislav, O. The Hxt13, Hxt15, Hxt16, and Hxt17 transporters of Saccharomyces cerevisiae represent a new type of polyol transporters. Scientific Reports; 2016, 6, 23502. |
| [47] | Barnett, J. A., Payne, R. W., Yarrow, D. Yeasts: Characteristics and Identification, 3rd edition. Cambridge University Press: 2000, p. 1139. |
| [48] | Antonio, C., Luis, C., Miguel, C., Felipe, S. Rapid isolation of mycoviral double-stranded RNA from Botrytis cinerea and Saccharomyces cerevisiae. Virology Journal; 2011, 8, 38, 1–7. |
| [49] | Andrea, C. O., María, E. R., Juan, M. O., Amparo, Q., Christian, A. L. Saccharomyces cerevisiae × Saccharomyces uvarum hybrids generated under different conditions share similar winemaking features. Wiley Online Library; 2018, 35, 157–171. |
APA Style
Fofana, B., Zoro, A. F., Toure, A., Kamagate, T., Traore, S., et al. (2026). Characterization of Fermentative Strains Isolated from Spontaneously Fermentation of Cashew Apples (Anacardium occidentale L.). International Journal of Microbiology and Biotechnology, 11(1), 9-19. https://doi.org/10.11648/j.ijmb.20261101.12
ACS Style
Fofana, B.; Zoro, A. F.; Toure, A.; Kamagate, T.; Traore, S., et al. Characterization of Fermentative Strains Isolated from Spontaneously Fermentation of Cashew Apples (Anacardium occidentale L.). Int. J. Microbiol. Biotechnol. 2026, 11(1), 9-19. doi: 10.11648/j.ijmb.20261101.12
@article{10.11648/j.ijmb.20261101.12,
author = {Bazoumana Fofana and Armel Fabrice Zoro and Abdoulaye Toure and Tidiane Kamagate and Safiatou Traore and Yade Rene Soro},
title = {Characterization of Fermentative Strains Isolated from Spontaneously Fermentation of Cashew Apples (Anacardium occidentale L.)},
journal = {International Journal of Microbiology and Biotechnology},
volume = {11},
number = {1},
pages = {9-19},
doi = {10.11648/j.ijmb.20261101.12},
url = {https://doi.org/10.11648/j.ijmb.20261101.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijmb.20261101.12},
abstract = {The cashew apple is a juicy and sweet fruit that loses its nutritional quality during the first week after falling from the cashew tree. This degradation is strongly linked to microorganisms that may present beneficial biotechnological potential. Therefore, the objective of this study is to characterize lactic acid bacteria and fermentative yeasts from cashew apples. Cashew apples collected from three localities (Tioro, Morovine, and Waraniene) in the Korhogo department (Northern Côte d’Ivoire) were washed in the laboratory, crushed, and subjected to fermentation. Samples were taken every two days over one week. Morphological studies focused on macroscopic and microscopic analyses of the strains. Physiological and biochemical tests of lactic acid bacteria and yeast isolates were all carried out in tubes. Molecular typing of these isolates was performed using the restriction enzymes DDeI or HinfI, and HaeIII. Characterization of lactic isolates revealed that all lactic acid bacteria isolated were bacillary in shape with catalase activity. They are homofermentative, mesophilic, acidophilic, and do not tolerate salinity beyond 4%. Moreover, they fermented lactose, glucose, maltose, fructose, starch, sucrose, and cellulose. They also produced exopolysaccharides. At the molecular level, all bands were observed at 1500 base pairs. Furthermore, the biochemical profile and enzymatic digestion performed with restriction enzymes identified the genus Lactobacillus, specifically Lactobacillus plantarum as the presumptive species. Characterization revealed yeasts of oval, elongated shape with lateral budding. Sediments were observed in their growth broths. They degraded urea and produced acetoin. They also fermented various sugars, except cellulose, citrate, and mannitol. All bands were observed at 500 base pairs. Biochemical tests and PCR confirmed that all isolates belonged to the genus Saccharomyces sp. Digestion with restriction enzymes (HaeIII and HinfI) divided them into two groups of Saccharomyces, of which 93.75% were Saccharomyces cerevisiae and 6.25% were Saccharomyces uvarum. The use of these presumptive species could serve in the production of cashew apple wine.},
year = {2026}
}
TY - JOUR T1 - Characterization of Fermentative Strains Isolated from Spontaneously Fermentation of Cashew Apples (Anacardium occidentale L.) AU - Bazoumana Fofana AU - Armel Fabrice Zoro AU - Abdoulaye Toure AU - Tidiane Kamagate AU - Safiatou Traore AU - Yade Rene Soro Y1 - 2026/02/06 PY - 2026 N1 - https://doi.org/10.11648/j.ijmb.20261101.12 DO - 10.11648/j.ijmb.20261101.12 T2 - International Journal of Microbiology and Biotechnology JF - International Journal of Microbiology and Biotechnology JO - International Journal of Microbiology and Biotechnology SP - 9 EP - 19 PB - Science Publishing Group SN - 2578-9686 UR - https://doi.org/10.11648/j.ijmb.20261101.12 AB - The cashew apple is a juicy and sweet fruit that loses its nutritional quality during the first week after falling from the cashew tree. This degradation is strongly linked to microorganisms that may present beneficial biotechnological potential. Therefore, the objective of this study is to characterize lactic acid bacteria and fermentative yeasts from cashew apples. Cashew apples collected from three localities (Tioro, Morovine, and Waraniene) in the Korhogo department (Northern Côte d’Ivoire) were washed in the laboratory, crushed, and subjected to fermentation. Samples were taken every two days over one week. Morphological studies focused on macroscopic and microscopic analyses of the strains. Physiological and biochemical tests of lactic acid bacteria and yeast isolates were all carried out in tubes. Molecular typing of these isolates was performed using the restriction enzymes DDeI or HinfI, and HaeIII. Characterization of lactic isolates revealed that all lactic acid bacteria isolated were bacillary in shape with catalase activity. They are homofermentative, mesophilic, acidophilic, and do not tolerate salinity beyond 4%. Moreover, they fermented lactose, glucose, maltose, fructose, starch, sucrose, and cellulose. They also produced exopolysaccharides. At the molecular level, all bands were observed at 1500 base pairs. Furthermore, the biochemical profile and enzymatic digestion performed with restriction enzymes identified the genus Lactobacillus, specifically Lactobacillus plantarum as the presumptive species. Characterization revealed yeasts of oval, elongated shape with lateral budding. Sediments were observed in their growth broths. They degraded urea and produced acetoin. They also fermented various sugars, except cellulose, citrate, and mannitol. All bands were observed at 500 base pairs. Biochemical tests and PCR confirmed that all isolates belonged to the genus Saccharomyces sp. Digestion with restriction enzymes (HaeIII and HinfI) divided them into two groups of Saccharomyces, of which 93.75% were Saccharomyces cerevisiae and 6.25% were Saccharomyces uvarum. The use of these presumptive species could serve in the production of cashew apple wine. VL - 11 IS - 1 ER -
Laboratory of Biotechnology and Valorization of Agroresources and Natural Substances, University Peleforo Gon Coulibaly, Korhogo, Côte d’Ivoire
Laboratory of Biotechnology and Valorization of Agroresources and Natural Substances, University Peleforo Gon Coulibaly, Korhogo, Côte d’Ivoire;Laboratory of Biotechnology, Felix Houphouet-Boigny University, Abidjan, Côte d’Ivoire
Laboratory of Biotechnology and Valorization of Agroresources and Natural Substances, University Peleforo Gon Coulibaly, Korhogo, Côte d’Ivoire;Laboratory of Biochemical Pharmacodynamy, University Felix Houphouet-Boigny, Abidjan, Côte d’Ivoire
Laboratory of Biotechnology and Valorization of Agroresources and Natural Substances, University Peleforo Gon Coulibaly, Korhogo, Côte d’Ivoire;Laboratory of Biochemical Pharmacodynamy, University Felix Houphouet-Boigny, Abidjan, Côte d’Ivoire
Laboratory of Biotechnology and Valorization of Agroresources and Natural Substances, University Peleforo Gon Coulibaly, Korhogo, Côte d’Ivoire

Figure 1. Crushed red and yellow cashew apples.
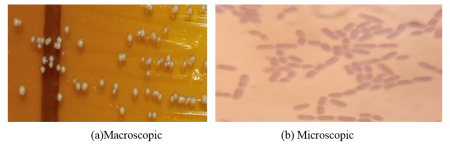
Figure 2. Appearance of Lactic Acid Bacteria.

Figure 3. Electrophoretic profile of the 16S gene and restriction digestion profiles of lactic acid bacterial genes using DDeI and HaeIII.
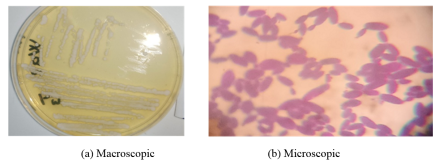
Figure 4. Appearance of yeasts.
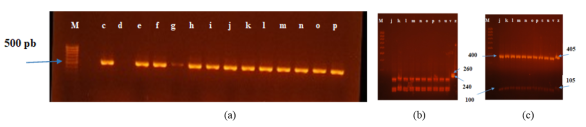
Figure 5. Electrophoretic profile of the 26S gene (a) and restriction digestion profiles of yeast genes using the enzymes HinfI (b) and HaeIII (c).

